Мускулы и нервные волокна, отвечающие за механизм мышечного сокращения, позволяют человеку совершать множество движений, а внутренним органам функционировать. В человеческом теле более 600 мышц, и все они различаются строением и физиологическими свойствами.
Определение
Мышечное сокращение – это механическая работа, которую производит мускул в результате процесса укорочения или развития напряженности.
Во время сокращения мышцы изменение длины вызывается тем, что тонкие волокна тянутся вдоль толстых волокон. Хотя длина перекрытия толстых и тонких нитей (саркомера) меняется, длина самих нитей остается прежней.
Возбуждение скелетных мышц, их произвольное сокращение стимулируется электрическими импульсами, переносимыми по нервам. Сокращения клеток сердечной мышцы стимулируются сердечной мускулатурой.
Существуют ещё непроизвольные сократительные мышечные движения. Они характерны для желудка, кишечника, мочевого пузыря, кровеносных сосудов, где сокращение гладкой мускулатуры вызывает перистальтику (сокращение волнообразное).
Типы
Механизм мышечного сокращения, физиология которого одинакова для всех типов мышц, состоит из процесса перехода мышцы из «состояния высокого напряжения» в «состояние низкого напряжения». 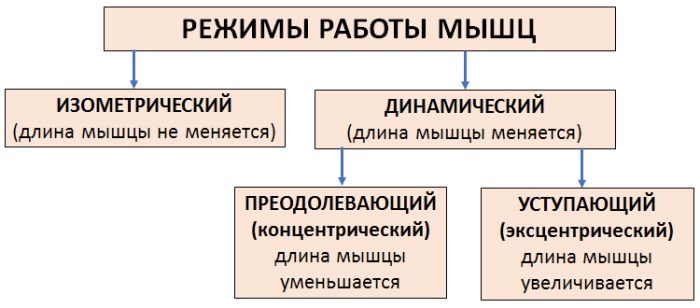 Иначе говоря, мышца напрягается, а затем снова расслабляется.
Иначе говоря, мышца напрягается, а затем снова расслабляется.
Изотонический режим
При этом виде сокращений мышца меняет длину, в то время как напряжение или сопротивление остаются прежними. В результате возникает движение части тела.
Существует два типа изотонического сокращения:
- Концентрическое. Это стандартное движение сокращения и подъема, когда мышца укорачивается и две точки ее соединения сближаются. При этом движение действует против силы тяжести и позволяет мышце тянуть и поднимать.
- Эксцентрическое сокращение задействует ту же мышцу, но действует на опускающуюся часть движения. По мере того, как мышца удлиняется, две точки соединения мышцы расходятся дальше, но мышца продолжает сокращаться и по-прежнему оказывает давление на вес. Оно действует как своего рода тормозной механизм, замедляя опускание под действием силы тяжести.
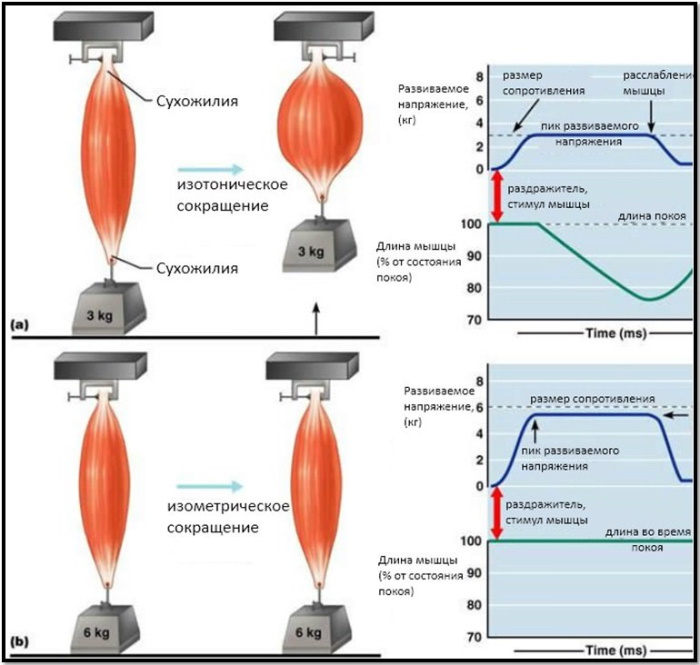 Эксцентрические сокращения противоположны концентрическим и возникают, когда мышца удлиняется при сокращении. Это можно наблюдать, например, при опускании гантели вниз в упражнении на сгибание бицепса. Мышца все еще сокращается, чтобы удерживать вес полностью, но двуглавая мышца удлиняется.
Эксцентрические сокращения противоположны концентрическим и возникают, когда мышца удлиняется при сокращении. Это можно наблюдать, например, при опускании гантели вниз в упражнении на сгибание бицепса. Мышца все еще сокращается, чтобы удерживать вес полностью, но двуглавая мышца удлиняется.
Изометрический режим
Механизм мышечного сокращения (физиология изометрического типа) в этом случае происходит без изменения мышечной длины. Изометрические сокращения отличаются от двух других типов тем, что не связаны ни с удлинением, ни с сокращением мышц. Скорее, это тип активации, при котором мышца сознательно напрягается, но связанные с ней суставы не двигаются.
Примеры включают в себя перенос объекта перед собой без движения, сидение на корточках у стены, не двигаясь, или удержание тела в положении равновесия в течение минуты или двух. В каждом случае мышца активируется без движения.
Это типично для мускул предплечий и кистей. Например, когда рука сжимает предмет, работают мышцы предплечий и кистей, но суставы руки не при этом не двигаются, а мышцы создают достаточную силу, чтобы не допустить падения предмета.
Другой пример — когда человек держит что-то в руке, в суставах при этом нет движения, но мышцы сокращаются, чтобы обеспечить силу, достаточную для удержания предмета.
Ауксотонический режим
Тип смешанного режима, в котором сочетаются элементы изотонического и изометрического сокращения относится к ауксотоническому типу. Этот вид характеризуется изменением тонуса и длины мышцы, имеет и другое название – динамический режим.
Для измерения такого типа сокращений требуется специальное оборудование, изокинетический динамометр. Изокинетические сокращения в повседневной жизни и при занятиях спортом встречаются редко. Лучше всего это можно наблюдать в режиме плавания брассом, когда вода обеспечивает постоянное, равномерное сопротивление движению приведения.
Виды
Характер мышечных сокращений зависит от порядка сокращений и частоты возникающих нервных стимулов.
Одиночное
В случае прямого раздражения мышцы или через нерв, который её иннервирует, возникает одиночное мускульное сокращение.
В этом механизме выделяют 3 фазы:
- скрытый период, от начала поступления импульса до ответа мускула, во время которого развивается потенциал действия (ПД);
- период сокращения или укорочения мышцы;
- период расслабления.
Мышечная возбудимость в этом случае меняется в соответствии с определенными фазами. Но в жизни одиночные импульсы человеку не поступают, а следуют друг за другом сериями, с определенными интервалами. На эти импульсы и отвечает мышца длительным сокращением.
Тетаническое
Когда продолжающееся сокращение мышц поддерживается непрерывно без расслабления, это называется тетаническим сокращением. Мышцы при этом могут укорачиваться, удлиняться или оставаться постоянной длины.
При этом движение стимулируется множественными импульсами с достаточно высокой частотой. Импульс, совпадающий с фазой расслабления способствует появлению целой серии последовательных одиночных сократительных движений.
Если частота импульсов повышается, то может произойти совмещение фазы расслабления предыдущего цикла с новым поступившим импульсом.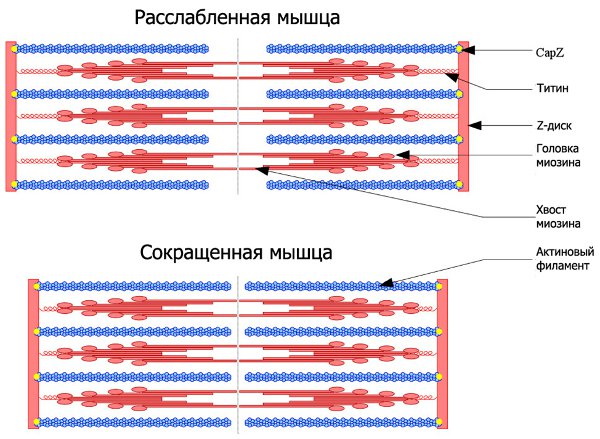
В таком случае говорят о зубчатом тетанусе, при котором происходит длительное сокращение, прерывающееся периодами неполного расслабления. Другим видом тетанического сокращения будет гладкий тетанус, когда длительное сокращение мышцы не прерывается периодами расслабления.
Тетаническое сокращение в более легкой форме является нормальным и происходит как процесс обычного физиологического явления во время таких действий, как, например, поднятие тяжести. В тяжелых случаях, при воспалительных и инфекционных состояниях, возникают тонические судороги симметричных групп мышц.
Сократительные белки миозина и актина
Отдельные мышечные клетки называются мышечными волокнами. Они длинные, цилиндрические и содержат несколько ядер. Эти мышечные волокна окружены клеточной мембраной, называемой сарколеммой. Внутри каждого мышечного волокна есть крошечные стержни, называемые миофибриллами, которые окружены саркоплазмой.
Миофибриллы состоят из повторяющихся сегментов, называемых саркомерами, которые представляют собой крошечные единицы, ответственные за сокращение скелетных мышц. Каждый саркомер содержит миофиламенты, белковые структуры из актина и миозина. Толстые миофиламенты состоят из миозина, а тонкие — из актина, тропонина и тропомиозина.
Рассмотрев структуру саркомера, можно заметить зигзагообразные участки, которые отмечают конечную точку каждого саркомера. Они называются Z-линиями, и участвуют в прикреплении тонких (актиновых) филаментов.
Миозин
Белок, толстая нить, выполняет структурную роль, являясь строительным блоком для толстых волокон, и функциональную, катализируя распад АТФ во время сокращения и взаимодействия с актином. 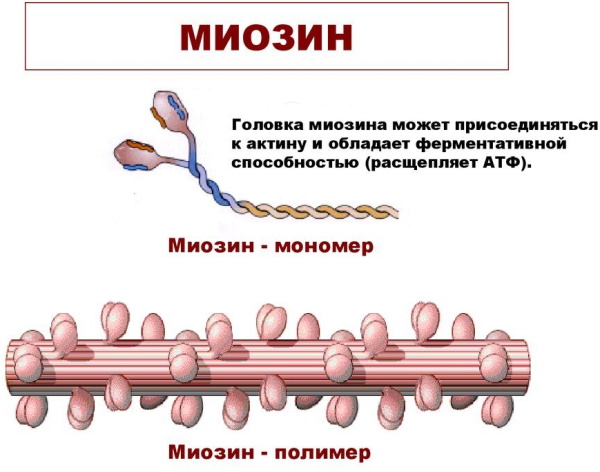 Отдельная молекула миозина имеет длину около 160 нм, асимметричную форму и содержит 2 основные белковые цепи.
Отдельная молекула миозина имеет длину около 160 нм, асимметричную форму и содержит 2 основные белковые цепи.
Актин
Актин – это основной компонент тонких волокон в мышцах, составляет около 25% белка миофиламентов. Отдельная клетка актина представляет собой единую белковую цепь, свернутую спиралью.
В мышце актин скручен в виде 2 длинных нитей бусообразных молекул. Тонкая нить актина состоит из чередующихся пучков, переплетенных с пучками толстых миозиновых нитей.
Тропомиозин
Тропомиозин — это длинный, спиралевидный белок, структурно похож на хвост молекулы миозина. Каждая молекула его контактирует с 7 единицами актина, охватывая большую часть актиновой нити. Молекула является регулятором мышечного сокращения, предотвращая прилипание миозина (и, следовательно, предотвращая сокращение). Внутри каждого белка тропомиозина находятся комплексы тропонина.
Тропонин
Тропонин состоит из комплекса белковых субъединиц. Одна часть белкового комплекса связана с молекулой тропомиозина, а другая – с ионами Ca2+. Тропонин, его молекула, расположена вдоль филамента через каждые 40 нм.
И тропонин, и тропомиозин являются регуляторами сокращающих и расслабляющих мышечных движений посредством связывания ионов Ca2+.
Преобразования
Физиологически человек – это биохимический двигатель, потому что пища — это топливо, которое расщепляется на составляющие: глюкозу (мельчайшие строительные блоки углеводов), аминокислоты и жиры. Механизм мышечного сокращения происходит за счёт использование глюкозы и энергии, обеспечиваемой АТФ.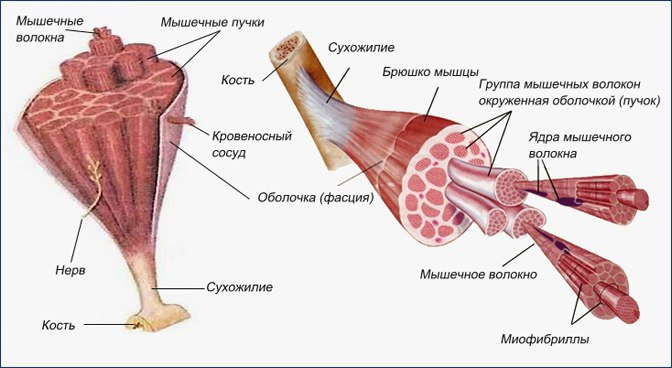
Скелетная мышца трансформирует химическую энергию в механическую. Таким образом процесс сокращения мышц происходит под воздействием 2-этапного молекулярного преобразования: электромеханического и хемомеханического.
Электрохимическое
Все живые клетки имеют мембраны с встроенными в него белками. Мембрана служит диффузионным барьером и изолятором для продвижения положительно и отрицательно заряженных ионов.
Нейроны и мышечные клетки через свои мембраны контролируют движение заряженных ионов с помощью избирательного открытия и закрытия ионных каналов. Таким образом образуются электрические токи и электросигналы. Хотя токи, создаваемые ионами, движущимися через канальные белки, очень малы, они составляют основу, как нейронной передачи сигналов, так и сокращения мышц.
Нервные импульсы электрически возбудимы, следовательно, они способны генерировать потенциал действия (ПД) — особый тип электрического сигнала, который перемещается по клеточной мембране в виде волны, быстро и точно передающей сигнал на большие расстояния.
Этот первый этап мышечного сокращения происходит за счёт возбуждения мембраны мышечного волокна, вдоль которой циркулирует потенциал действия. Это свойство называется возбудимостью, когда ПД генерирует нейроны и мышечные клетки.
Хемомеханическое
Сократительная работа мышц происходит за счет выделяемой в процессе окисления углеводов или липидов энергии.
В процессе механохимической реакции происходит поэтапное преобразование химической энергии в механическую:
- вначале идет процесс связывания тропонина с ионами Ca2+;
- следующий шаг – взаимодействие головки тяжелой цепи миозина с актином;
- происходит скольжение актина и миозина относительно друг друга,
Этапы сокращений
Бегает человек, едет на велосипеде или поднимает тяжести, все движения являются частями сложного процесса координации мышечных сокращений.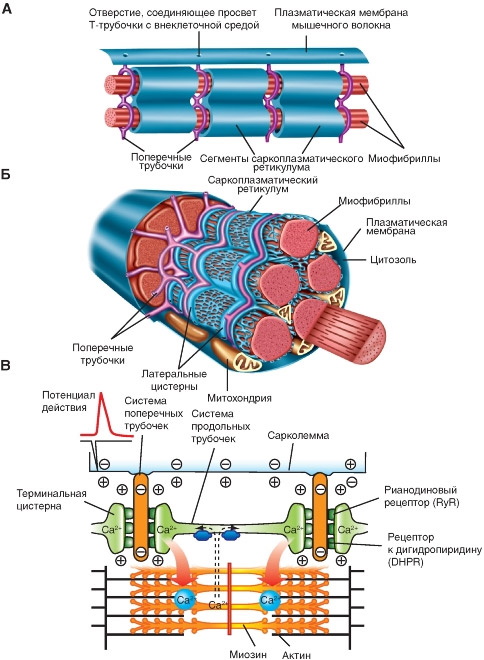
В состоянии покоя ионы Са2+, участвующие в процессе мышечных сокращений, хранятся в цитоплазме клетки, или саркоплазматическом ретикулуме. При этом не возникает связь возбуждения-сокращения, которая преобразует электрический сигнал нейрона через ацетилхолин в электросигнал на мышечной мембране.
Механизм мышечного сокращения, физиология которого зависит от свойства скелетных мышц, происходит только при определенной последовательности событий.
Сократительная работа начинается, когда саркомеры укорачиваются, толстые и тонкие волокна скользят друг мимо друга, по принципу скольжения нитей. Белки при этом, отвечающие за чувствительность мышц к ионам Ca2+, такие как тропонин и тропомиозин, контролируют образование поперечных мостиков, появляющихся в процессе взаимодействия актина и миозина.
| Этапы мышечного сокращения | Действия |
| I | Происходит процесс присоединения кальция к тропонину. |
| II | Начинается сцепление миозинового мостика с актином. |
| III | Идёт циклическое движение поперечного мостика. |
| IV | Происходит диссоциация и образование комплекса между миозином и актином. |
| V | Поворот головки миозинового поперечного мостика. |
| VI | Завершение цикла при распаде АТФ. |
Процесс сокращения мышц длится до тех пор, пока имеются необходимые запасы ионов Са2+ и энергии (АТФ). Как только импульс прекращается, ионы возвращаются обратно в саркоплазматический ретикулум, актин переходит в состояние покоя, а мышцы удлиняются и расслабляются.
Первый
Сигнал к мышечному сокращению исходит от нейронов, передается в точку контакта двигательного нерва с мышцей. Или, другими словами, электрический импульс проходит от тела двигательной нервной клетки в спинном мозге вдоль нервного аксона к месту назначения, нервно-мышечному соединению.
Второй
Электрический импульс, вызывающий деполяризацию и ПД в сарколемме, запускает процесс высвобождения ионов кальция из саркоплазматического ретикулума. Затем происходит взаимодействие ионов кальция с комплексом тропонина на актиновом миофиламенте. Это даёт толчок к смещению комплекса тропонина и тропомиозина, что приводит к разблокировке места связывания миозина с молекулами актина.
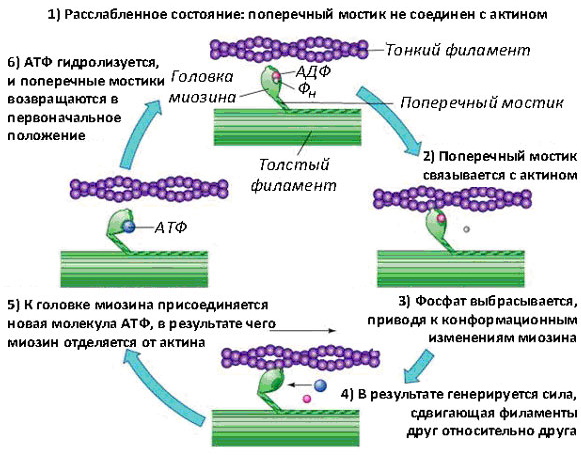
В момент, когда сигнал для сокращения направляется по нерву к мышечной клетке, происходит активация белков, актина и миозина. Миозин начинает работать как двигатель, высвобождая энергию АТФ, что способствует скольжению миозиновой нити вдоль актиновой.
В каждой молекуле акттина имеется активный центр, который перекрыт белками, тропонином и тропомиозином. Они не позволяют раньше времени вступать в действие актину с миозином.
Как только место связывания миозина обнажается, головка этой молекулы образует соединение с актиновым филаментом. В этом случае говорят о том, что миофиламент актина находится в стадии «включён».
Третий
После «включения» актина миозиновая головка сгибается в шарнирных областях, прикрепляясь к соседнему актиновому филаменту. При этом образуется активный поперечный мостик. Он работает как фермент (миозин-АТФ), который расщепляет запасенный аденозинтрифосфат (АТФ), сохраняющийся в головке миозина, в аденозиндифосфат (АДФ) и неорганический фосфат, высвобождая при этом энергию.
Четвертый
Энергия, освободившаяся при гидролизе, используется для движения миозиновой головки относительно нити актина. Головка тяжёлой цепи миозина наклоняется и начинает тянуть актиновую нить, при этом происходит движение толстых и тонких нитей друг относительно друга.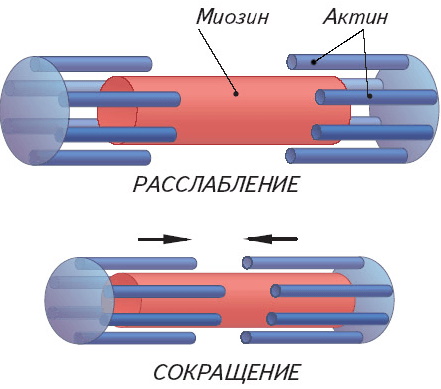 Противоположные концы актиновых миофиламентов внутри саркомера начинают встречное движение, что приводит к сократительному движению мышц.
Противоположные концы актиновых миофиламентов внутри саркомера начинают встречное движение, что приводит к сократительному движению мышц.
Пятый
Механизм мышечного сокращения (физиология сложного процесса многошаговая) на этом этапе заключается в том, что головная часть цепи миозина, используя энергию гидролиза АТФ, прикрепляется к участку воздействия актина. Высвобождение неорганического фосфата усиливает связывающее взаимодействие между миозином и актином и впоследствии запускает «силовой удар».
Силовой удар — это ключевой этап создания силы, используемый моторными белками миозина. Силы генерируются на актиновой нити, когда белок миозина возвращается к своей исходной форме. Актиновые нити вытягиваются. Именно на этом этапе происходит сокращение мышцы.
Шестой
Когда миозин восстанавливает свою первоначальную форму или конформацию, АДФ высвобождается, но головка миозина остается плотно связанной с нитью в новом месте, тем самым возвращая цикл к началу. Распад АТФ от миозина завершает цикл, и весь комплекс актин-миозин остается в фиксированном состоянии.
Миозиновая головка продолжает занимать позицию под углом в 45˚к тонким и толстым нитям. В этих условиях большое количество головок миозина, свободных от АДФ остается связанными с актином до тех пор, пока новая молекула АТФ не свяжется с ним и тем самым начнет новый сократительный цикл.
Один нейронный удар приводит только к укорочению примерно на 1% всей мышцы. Поэтому для достижения общего сокращения до 35%, весь процесс необходимо повторять много раз. Считается, что в то время как половина поперечных мостиков активна в протягивании актина через миозин, другая половина ищет свой следующий путь связывания.
Для живой системы характерен механизм мышечного сокращения за счёт взаимодействия белков. Сократимость – это свойство клеток любых тканей. Но только мышечные имеют сложную физиологическую структуру, которая приводит к разнообразным движениям.
Видео о мышечных сокращениях
Физиология. Механизмы мышечного сокращения: